📋 Key Information Summary
- Natural killer (NK) cells are innate lymphoid cells (ILC1 group) comprising 5–15% of peripheral blood lymphocytes that kill target cells without prior sensitisation.
- NK cell activation is governed by a balance between activating receptors (NKG2D, NKp46, NKp30, DNAM-1) and inhibitory receptors (KIR, NKG2A/CD94, LILRB1) that recognise MHC class I.
- The "missing-self" hypothesis explains NK cell cytotoxicity: downregulation of MHC class I on infected or transformed cells removes inhibitory signals and triggers killing.
- Cytotoxic mechanisms include perforin/granzyme release (primary), death receptor signalling (FasL, TRAIL), and antibody-dependent cellular cytotoxicity (ADCC) via CD16 (FcγRIIIa).
- Clinically relevant in haematopoietic stem cell transplantation (HSCT), adoptive NK cell immunotherapy, and solid organ transplant rejection monitoring.
- NK cell deficiency syndromes are rare primary immunodeficiencies presenting with severe, recurrent herpesvirus infections (CMV, EBV, HSV, VZV).
- Killer immunoglobulin-like receptor (KIR)–HLA mismatch in haploidentical HSCT is associated with reduced relapse and graft-versus-host disease (GvHD) in acute leukaemia.
- Anti-CD20 monoclonal antibodies (rituximab, obinutuzumab) rely on NK cell–mediated ADCC as a primary mechanism of action.
- NK cell function testing (flow cytometry–based cytotoxicity assay, CD107a degranulation) is available at major Australian immunology reference laboratories (RCH Melbourne, Westmead).
- Low NK cell activity has been associated with recurrent pregnancy loss, though routine NK cell testing in reproductive medicine remains controversial per RANZCOG guidance.
- Australian research programmes at Peter MacCallum Cancer Centre and QIMR Berghofer are pioneering CAR-NK cell therapies for haematological malignancies.
Introduction & Australian Epidemiology
Natural killer (NK) cells are innate lymphoid cells (ILC1-type) that constitute approximately 5–15% of circulating peripheral blood lymphocytes in healthy adults. Unlike T and B lymphocytes, NK cells do not require antigen-specific receptor gene rearrangement and provide rapid cytotoxic responses against virally infected cells and tumour cells without prior sensitisation.
NK cells were first described in 1975 by researchers at the Karolinska Institute and the National Cancer Institute, identified by their spontaneous ability to lyse tumour cells in vitro. They are now recognised as critical effectors bridging innate and adaptive immunity, producing cytokines (IFN-γ, TNF-α, GM-CSF) that shape downstream immune responses.
In the Australian context, NK cell biology is relevant across several clinical domains:
- Haematopoietic stem cell transplantation: Australian transplant centres (Royal Adelaide Hospital, Westmead Hospital, Peter MacCallum Cancer Centre) utilise KIR-ligand mismatch models in haploidentical HSCT protocols.
- Primary immunodeficiency: Approximately 1 in 500,000 births result in classical NK cell deficiency syndromes; the Australian Paediatric Immunodeficiency Consortium tracks these rare diagnoses.
- Immunotherapy development: Australian institutions participate in CAR-NK cell clinical trials, with active programmes at QIMR Berghofer Medical Research Institute and the Peter MacCallum Cancer Centre.
- Reproductive medicine: Peripheral blood NK cell testing is frequently requested in Australian fertility clinics despite lack of endorsement by RANZCOG, generating significant clinical debate.
- Solid tumour immunology: Tumour-infiltrating NK cells are prognostic biomarkers in several malignancies managed in Australian oncology practice.
NK Cell Receptors
NK cell function is regulated by a repertoire of germline-encoded activating and inhibitory surface receptors. The integration of signals from these receptors determines whether a target cell is killed or spared. Unlike T-cell receptors, NK cell receptors are not generated by somatic recombination and are encoded within the leukocyte receptor complex (LRC) on chromosome 19q13.4 and the natural killer gene complex (NKC) on chromosome 12p13.
Inhibitory Receptors
| Receptor | Ligand | Gene Complex | Clinical Significance |
|---|---|---|---|
| KIR2DL1 (CD158a) | HLA-C2 (Lys80) | LRC (19q13.4) | HSCT mismatch model; preeclampsia association |
| KIR2DL2/3 (CD158b) | HLA-C1 (Asn80) | LRC (19q13.4) | HSV-1 susceptibility; centipede allergy |
| KIR3DL1 (CD158e1) | HLA-Bw4 | LRC (19q13.4) | HIV progression; HSCT outcomes |
| KIR3DL2 (CD158k) | HLA-A3/A11; free HLA-B27 dimer | LRC (19q13.4) | Spondyloarthritis pathogenesis |
| NKG2A/CD94 | HLA-E | NKC (12p13) | Therapeutic target (monalizumab, anti-NKG2A mAb) |
| LILRB1 (ILT2/LIR-1) | Broad HLA class I | LRC (19q13.4) | CMV immune evasion via UL18 homologue |
| TIGIT | CD155 (PVR), CD112 (Nectin-2) | — | Checkpoint inhibition target; NK exhaustion |
Activating Receptors
| Receptor | Ligand(s) | Signalling Adaptor | Function |
|---|---|---|---|
| NKG2D | MICA, MICB, ULBP1–6 | DAP10 | Stress-induced ligand recognition; tumour immunosurveillance |
| NKp46 (NCR1) | Viral haemagglutinins; tumour ligands (unknown) | CD3ζ, FcεRIγ | Primary natural cytotoxicity receptor; influenza recognition |
| NKp30 (NCR3) | B7-H6, BAG6 | CD3ζ, FcεRIγ | DC cross-talk; CMV immune evasion (pp65 inhibition) |
| NKp44 (NCR2) | Viral haemagglutinins; proliferating cell nuclear antigen (PCNA) isoforms | DAP12 | Expressed only on activated NK cells |
| DNAM-1 (CD226) | CD155 (PVR), CD112 (Nectin-2) | — | Tumour recognition; competes with TIGIT |
| CD16 (FcγRIIIa) | IgG Fc (opsonised targets) | CD3ζ, FcεRIγ | ADCC; therapeutic antibody mechanism |
KIR Genetics and HLA Epitope Matching
KIR genes are highly polymorphic and organised in haplotypes. The two major haplotype groups are:
- KIR-A haplotype: Contains mainly inhibitory KIR genes (2DL1, 2DL3, 3DL1, 3DL2, 3DL3) with a single activating gene (2DS4). Associated with reproductive success and susceptibility to autoimmune disease.
- KIR-B haplotype: Contains additional activating KIR genes (2DS1, 2DS2, 2DS3, 2DS5, 3DS1). Associated with improved outcomes in haploidentical HSCT and protection against CMV reactivation.
KIR typing is performed by PCR-SSP or next-generation sequencing at specialised Australian HLA laboratories including the Australian Red Cross Lifeblood HLA laboratory and state-level tissue typing services.
Activation Mechanisms
NK cell activation is determined by the net balance of signals received from activating and inhibitory receptors. Several models describe how this signal integration occurs:
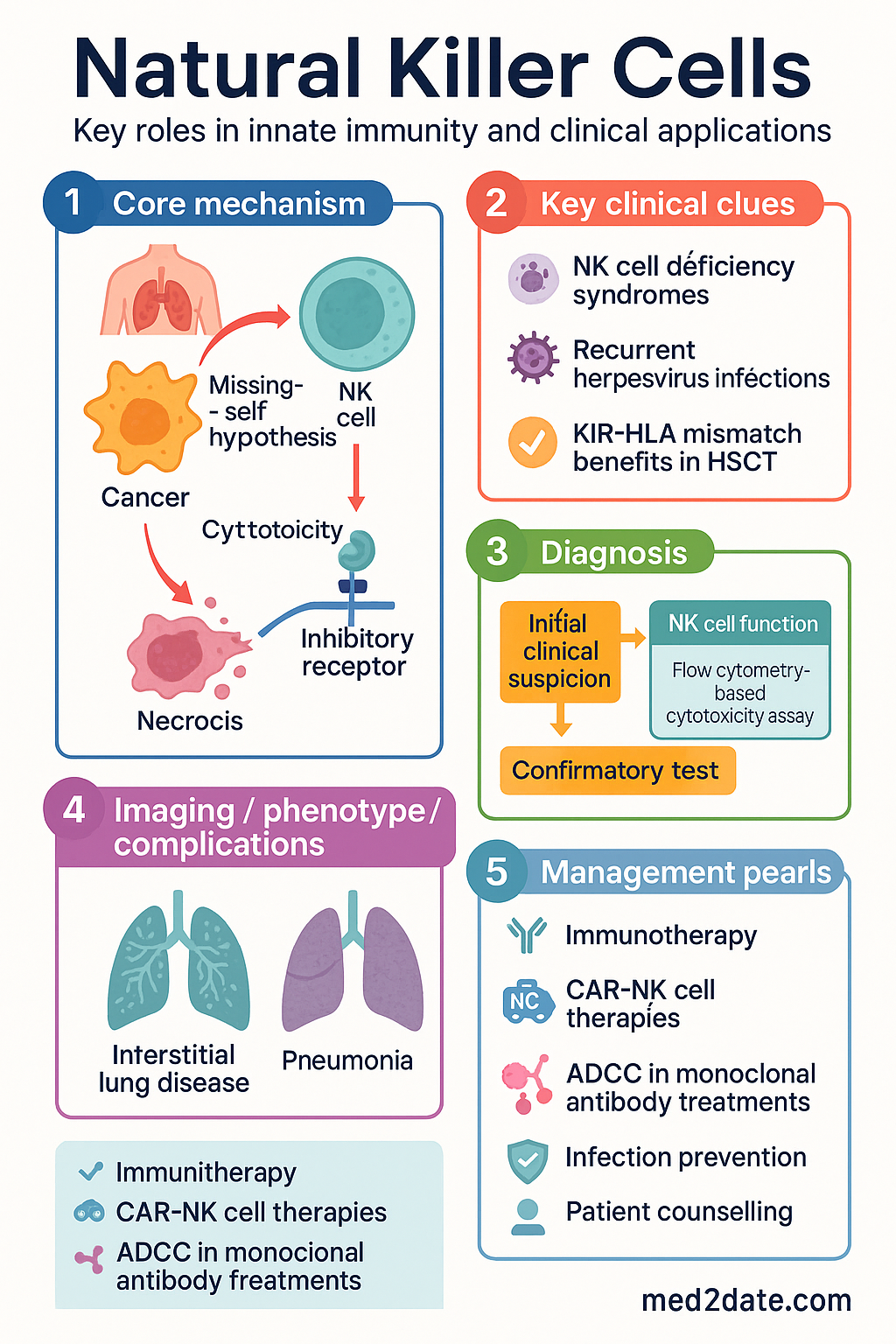
The Missing-Self Hypothesis
The foundational model of NK cell biology proposes that NK cells survey target cells for expression of MHC class I molecules. Normal cells expressing adequate MHC class I engage inhibitory KIR receptors and are spared. Cells that downregulate MHC class I — a common immune evasion strategy during viral infection or malignant transformation — lose inhibitory signalling and become susceptible to NK-mediated killing.
The Induced-Self Model
Stress, infection, or transformation induces expression of ligands for activating receptors (particularly NKG2D ligands: MICA, MICB, ULBP1–6) on target cells. These stress-induced ligands overcome inhibitory signals and trigger NK cell activation even when MHC class I expression is maintained. NKG2D ligands are upregulated by DNA damage response pathways, heat shock, and viral infection.
The Non-Self Model
Some NK cell receptors directly recognise pathogen-derived ligands. For example, NKp46 recognises influenza haemagglutinin and Sendai virus haemagglutinin-neuraminidase. This provides direct pathogen detection analogous to Toll-like receptor signalling.
Licensing and Education
Not all NK cells are equally functional. The process of NK cell education (also termed "licensing" or "arming") ensures self-tolerance:
- Licensing model: NK cells that express inhibitory receptors capable of recognising self-MHC class I during development become functionally competent ("licensed"). NK cells lacking such interactions remain hypo-responsive ("unlicensed").
- Disarming model: NK cells that receive chronic activating signals in the absence of inhibitory input become anergised to prevent autoimmunity.
- Rheostat model: NK cell responsiveness is tuned quantitatively according to the cumulative strength of inhibitory receptor engagement during development, rather than a binary licensed/unlicensed state.
Cytokine-Mediated Activation
NK cells are potentiated by several cytokines that enhance cytotoxicity, proliferation, and cytokine production:
Cytotoxic Mechanisms
NK cells kill target cells through three principal mechanisms. These pathways are utilised in both physiological immunosurveillance and therapeutic settings such as antibody therapy and adoptive cell transfer.
1. Perforin–Granzyme Pathway (Primary)
The dominant cytotoxic mechanism involves directed secretion of cytolytic granules containing perforin and granzymes:
- Immunological synapse formation: NK cell engages target cell, forming a tight intercellular junction. LFA-1 and CD2 on the NK cell bind ICAM-1 and CD58 on the target.
- Cytoskeletal polarisation: The microtubule-organising centre (MTOC) and lytic granules polarise toward the synapse.
- Granule exocytosis: Granules fuse with the NK cell membrane and release contents into the synaptic cleft.
- Pore formation: Perforin monomers insert into the target cell membrane and oligomerise to form transmembrane pores (analogous to complement C9/MAC).
- Granzyme entry and apoptosis: Granzyme B enters through perforin pores (or via mannose-6-phosphate receptor endocytosis), cleaves caspase-3, caspase-7, and BID, triggering apoptotic cascades.
2. Death Receptor Pathway
NK cells express death receptor ligands that trigger apoptosis in target cells through extrinsic signalling:
- Fas ligand (FasL/CD178): Binds Fas (CD95) on target cells, recruiting FADD and activating caspase-8 → caspase-3 cascade.
- TRAIL (TNF-related apoptosis-inducing ligand): Binds TRAIL-R1 (DR4) and TRAIL-R2 (DR5), activating the same caspase cascade. Preferentially kills transformed cells over normal cells — basis for ongoing therapeutic interest.
- TNF-α: Can induce apoptosis via TNF-R1 in certain contexts, though primarily pro-inflammatory.
3. Antibody-Dependent Cellular Cytotoxicity (ADCC)
CD16 (FcγRIIIa) on NK cells binds the Fc region of IgG antibodies coating target cells, triggering degranulation without requirement for additional activating signals. This mechanism is central to the efficacy of several therapeutic monoclonal antibodies used in Australian oncology practice:
| Antibody | Target | Indication | ADCC Contribution | PBS Status |
|---|---|---|---|---|
| Rituximab | CD20 | NHL, CLL, RA, ANCA vasculitis | Major mechanism | ✔ PBS |
| Obinutuzumab | CD20 (glycoengineered) | CLL, FL | Enhanced ADCC (afucosylated Fc) | Restricted |
| Trastuzumab | HER2 | Breast, gastric cancer | Significant contributor | ✔ PBS |
| Cetuximab | EGFR | CRC, HNSCC | Contributor; FcγRIIIa polymorphism affects response | Restricted |
| Daratumumab | CD38 | Multiple myeloma | Major mechanism | Restricted |
Cytokine Secretion
Beyond direct cytotoxicity, NK cells produce cytokines that modulate the broader immune response:
- IFN-γ: Activates macrophages, upregulates MHC expression, promotes Th1 polarisation. Critical for defence against intracellular pathogens.
- TNF-α: Pro-inflammatory; enhances endothelial activation and neutrophil recruitment.
- GM-CSF: Stimulates myeloid cell differentiation and dendritic cell maturation.
- Chemokines (CCL3, CCL4, CCL5, XCL1): Recruit additional immune cells to sites of infection or tumour.
Clinical Relevance
NK cell biology intersects clinical medicine across haematology, immunology, oncology, transplantation, and reproductive medicine. This section covers major clinical applications and disease associations relevant to Australian practice.
NK Cell Deficiency Syndromes
Primary NK cell deficiencies are rare inborn errors of immunity classified by the International Union of Immunological Societies (IUIS). They present with selective susceptibility to herpesvirus infections:
| Condition | Gene | NK Cell Phenotype | Key Clinical Features |
|---|---|---|---|
| NKD (classical NK deficiency) | GATA2 (haploinsufficiency), MCM4 | Absent or very low NK cells | Severe CMV, HSV, EBV; myelodysplasia (GATA2) |
| Functional NK deficiency | MCM4, FCGR3A (CD16), IRF8 | Normal count, impaired cytotoxicity | HSV encephalitis, severe varicella, EBV lymphoma |
| XLP1 (Duncan syndrome) | SH2D1A (SAP) | Impaired NKT cells; variable NK dysfunction | EBV-driven haemophagocytic lymphohistiocytosis, lymphoma |
| FHL2 (perforin deficiency) | PRF1 | Present but non-cytotoxic | Infantile HLH, >50% mortality without HSCT |
Australian diagnostic pathway: Suspect NK cell deficiency in patients with severe, recurrent, or disseminated herpesvirus infections. Initial testing includes:
- Full blood count with lymphocyte subset analysis (CD3, CD4, CD8, CD16+CD56+ NK cells) — MBS item 69487
- NK cell functional assay (CD107a degranulation, flow cytometry–based cytotoxicity) — available at Royal Children's Hospital Melbourne, Westmead Hospital
- Perforin expression by flow cytometry (screening for FHL2)
- Genetic panel or whole-exome sequencing via approved genetics services (subject to Medicare genomic testing criteria from November 2024)
Haematopoietic Stem Cell Transplantation
KIR-ligand mismatch between donor and recipient is exploited therapeutically in haploidentical HSCT. The "graft-versus-leukaemia" (GvL) effect mediated by alloreactive NK cells has been demonstrated most convincingly in acute myeloid leukaemia (AML):
Australian HSCT centres performing KIR-based donor selection include Royal Adelaide Hospital, Westmead Hospital (Sydney), Peter MacCallum Cancer Centre (Melbourne), and Royal Brisbane and Women's Hospital.
Adoptive NK Cell Immunotherapy
Adoptive transfer of expanded NK cells is an evolving therapeutic modality. Current approaches include:
- Unmodified NK cells: Expanded ex vivo from peripheral blood, umbilical cord blood, or NK-92 cell line with IL-15 or IL-2. Used in relapsed AML, high-risk MDS.
- CAR-NK cells: NK cells engineered with chimeric antigen receptors (e.g., anti-CD19 CAR-NK for B-lymphoid malignancies). Early data show comparable efficacy to CAR-T with lower rates of cytokine release syndrome (CRS) and immune effector cell-associated neurotoxicity syndrome (ICANS).
- Memory-like NK cells: Brief pre-activation with IL-12/15/18 generates long-lived NK cells with enhanced recall responses. Phase I/II trials in AML show promising remission rates.
NK Cells in Solid Organ Transplantation
The role of NK cells in solid organ transplant rejection is increasingly recognised:
- Antibody-mediated rejection (AMR): NK cells contribute to endothelial injury through ADCC via CD16 engagement with donor-specific antibodies (DSA). Tissue transcriptomic studies show NK cell–associated transcripts (GNLY, PRF1, CXCL10) in kidney biopsies with AMR.
- T cell–mediated rejection: NK cells may amplify rejection through IFN-γ production and dendritic cell activation.
- Tolerance: Some studies of operational tolerance in kidney transplant recipients show enrichment of NK cell gene signatures, suggesting a potential tolerogenic role.
Reproductive Medicine
Uterine NK (uNK) cells are the predominant leucocyte population in the decidua during early pregnancy (comprising 70% of decidual immune cells). They are phenotypically distinct from peripheral blood NK cells (CD56bright CD16− rather than CD56dim CD16+) and function in spiral artery remodelling and trophoblast invasion rather than cytotoxicity.
NK Cells in Viral Infection
NK cells are critical early responders to viral infection, particularly herpesviruses:
- CMV: Drives expansion of NKG2C+ CD57+ adaptive NK cells with enhanced ADCC. CMV seropositivity is associated with marked NK cell repertoire changes persisting lifelong. CMV encodes multiple immune evasion proteins (UL16, UL18, UL40, UL83/pp65) targeting NK cell recognition.
- EBV: NK cells control primary EBV infection. Patients with XLP1 or NK cell deficiencies develop life-threatening EBV-driven lymphoproliferation and HLH.
- HIV: NK cells contribute to viral control; KIR3DL1 + HLA-Bw4-80I combination is associated with slower progression to AIDS.
- Influenza: NKp46 directly recognises influenza haemagglutinin. NK cell–deficient mice show increased mortality from influenza challenge.
- SARS-CoV-2: Severe COVID-19 is associated with NK cell exhaustion, reduced NKG2A/NKG2D expression, and lymphopenia. Therapeutic IL-15 administration is under investigation to restore NK cell function.
Malignancy and Tumour Immunosurveillance
NK cells provide immunosurveillance against malignancy through recognition of stress ligands and loss of MHC class I:
- Tumour escape mechanisms: TGF-β secretion, shedding of soluble MICA/MICB (sMIC), hypoxia, and IDO-mediated tryptophan depletion suppress NK cell function in the tumour microenvironment.
- Checkpoint blockade: Anti-NKG2A (monalizumab), anti-TIGIT (tiragolumab), and anti-KIR (lirilumab) antibodies are in clinical trials as combination strategies with anti-PD-1/PD-L1 therapy.
- Prognostic significance: High tumour-infiltrating NK cell density correlates with improved survival in colorectal, gastric, and hepatocellular carcinomas. Intratumoral NK cell assessment is being incorporated into pathological scoring systems.
Investigations
NK cell assessment requires specialist immunology laboratory facilities. The following tests are available in Australia:
Special Populations
Aboriginal and Torres Strait Islander Health
📚 References
- 1. Caligiuri MA. Human natural killer cells. Blood. 2008;112(3):461–469. doi:10.1182/blood-2007-09-077438
- 2. Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nature Immunology. 2008;9(5):503–510. doi:10.1038/ni1582
- 3. Bhatt DL, Mena-Hurtado C, et al. NK cells and cancer immunosurveillance. Nature Reviews Immunology. 2018;18(8):491–504.
- 4. Ruggeri L, Capanni M, Urbani E, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science. 2002;295(5562):2097–2100. doi:10.1126/science.1068440
- 5. Mace EM, Bigley V, Gunesch JT, et al. Biallelic mutations in IRF8 impair human NK cell maturation and function. Journal of Clinical Investigation. 2017;127(1):306–320. doi:10.1172/JCI86276
- 6. Cooper MA, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends in Immunology. 2001;22(11):633–640. doi:10.1016/S1471-4906(01)02060-9
- 7. Liu LL, Béziat V, Oei VYS, et al. Ex vivo expanded adaptive NK cells effectively kill primary acute lymphoblastic leukemia cells. Cancer Immunology Research. 2017;5(8):654–665. doi:10.1158/2326-6066.CIR-16-0296
- 8. Hsu KC, Keever-Taylor CA, Wilton A, et al. Improved outcome in HLA-identical sibling hematopoietic stem-cell transplantation for acute myelogenous leukemia predicted by KIR and HLA genotypes. Blood. 2005;105(12):4878–4884. doi:10.1182/blood-2004-12-4825
- 9. Gleeson M, Clancy RL, Cripps AW, et al. Mucosal immune response to respiratory infections in Australian Indigenous populations. Immunology and Cell Biology. 2003;81(4):267–272.
- 10. RANZCOG. Recurrent Pregnancy Loss. College Statement (C-Obs 42). Royal Australian and New Zealand College of Obstetricians and Gynaecologists; 2021.
- 11. American Society for Reproductive Medicine (ASRM). Evaluation and treatment of recurrent pregnancy loss: a committee opinion. Fertility and Sterility. 2012;98(5):1103–1111. doi:10.1016/j.fertnstert.2012.06.048
- 12. Orange JS. Natural killer cell deficiency. Journal of Allergy and Clinical Immunology. 2013;132(3):515–525. doi:10.1016/j.jaci.2013.07.020
- 13. Cichocki F, Felices M, McCullar V, et al. Cutting edge: microRNA-181 promotes human NK cell development by regulating Notch signaling. Journal of Immunology. 2011;187(12):6171–6175. doi:10.4049/jimmunol.1100835
- 14. Australian Institute of Health and Welfare (AIHW). Aboriginal and Torres Strait Islander Health Performance Framework 2020 summary report. Canberra: AIHW; 2020.
- 15. Lanier LL. NK cell recognition. Annual Review of Immunology. 2005;23:225–274. doi:10.1146/annurev.immunol.23.021704.115526